
Indução
Antes da fecundação, a orientação do ovo é aleatória, apenas uma meia-hora depois da fusão das membranas plasmáticas do espermatozóide e do ovo é que este se oriente com o polo animal em cima e o vegetativo em baixo numa rotação de cerca de 30°. A rotação de 30° orienta os organelos, as proteínas e o RNAm e estabelece a região dorsal, ou seja, é fundamental ao desenvolvimento correcto do embrião .

Fig.1 - Diferentes rotações do ovo e consequências no embrião
A zona de entrada do espermatozóide determina sempre a parte ventral do ovócito e a parte oposta é a parte dorsal.
A determinação do eixo antero-posterior: formação do cérebro na parte anterior, da medula espinal na parte posterior e da mesoderme paraxial no centro (que vai dar os somitos).
A indução é o processo pelo qual uma célula, um grupo de células ou um tecido indutores envolvem uma célula, um grupo de células ou um tecido indutivos numa nova via de diferenciação.

Fig.2 - Indução da mudança de forma de uma célula por sinais de outras células
A indução da mesoderme é a primeira que existe. As células vegetativas (do polo vegetativo) que correspondem à endoderme vão enviar sinais às células animais (do polo animal) que correspondem à ectoderme que vão passar a ser mesodérmicas.
As células não enviam todas os mesmos sinais, depende se são da parte ventral ou dorsal - indução regional.
As células da parte dorsal enviam os sinais do centro de Nieuwkoop para uma zona chamada de organizador.

Fig.3 - Envio de sinais mesodérmicos dorsais

Fig.4 - Experiências de Dale e Slack, 1987. Diferentes grupos de células vegetais provocam diferentes tipos de induções: dorsais, intermédias ou ventrais

Fig.5 - Experiências de Gimlich e Gerhart, 1984 e 1986. Transplantação do centro de Nieuwkoop para a região ventral de outro embrião e consequências nesse mesmo embrião.
A indução pode ser feita de três maneiras diferentes:

Fig.6 - As diferentes maneiras de indução
O fenómeno de dorsalização é influenciado pela região dorsal sobre região ventral, ou seja, a mesoderme dorsal influencia o destino da mesoderme ventral.
Moléculas indutoras e moléculas modificadoras:
- FGFs (Fibroblast growth factor)
- transcritos maternais
- blastoméros vegetativos
- mantêm a indução e diferenciação da mesoderme
- TGFs (Transforming growth factors)
- Activina - actua na morfogénese bloqueando os FGFs e os seus efeitos variam conforme a sua concentração.
- Vg1 - é sintetizada depois da fecundação sob uma forma inactiva. É activada pela rotação cortical na região dorsal.
- BMPs
- Nodal - presente sob um gradiente de concentração e temporal
- Factores de transcrição
- siamois
- proteína Vegt.
- Dorsalização e ventralização
- BMP4 e Xwnt8
- Noggin, Chordin
No início da fecundação a proteína Dsh está localizada no polo vegetativo. Imediatamente após a rotação cortical, a proteína Dsh (disheveld) está posicionada na região dorsal.
A proteína GSK3 é inibida na região dorsal pela Dsh, mas activada na região ventral. Depois, esta proteína vai degradar beta-catenina na região ventral então esta encontra-se apenas na região dorsal onde se acumula e é estável. As moléculas de beta-catenina vão activar a transcrição do gene siamois, mas a proteína GSK3 pode inibir a molécula beta-catenina e não haverá dorsalização.

Fig.7 - Modo de acção das proteínas Dsh e GSK3

Fig.8 - Na região ventral não vai haver transcrição do gene siamois como na região dorsal

Fig.9 - Modo de acção e localização de outras moléculas indutoras e modificadoras
Indução neural
As células da mesoderme posicionam-se progressivamente na ectoderme na região dorsal do embrião. Estas células induzem o estabelecimento da placa neural e depois do tubo neural a partir da ectoderme (indução das células da endoderme por contacto com as células da mesoderme).
O lábio dorsal do blastoporo induz a formação do tubo neural, então o tubo neural e os seus derivados são formados a partir da ectoderme.
Organisador de Spemann:
- centro indutor
- indutor da gastrulação
- dorsalização da mesoderme
O contacto directo da mesoderme com a ectoderme permite a indução e a formação do tubo neural na parte dorsal.
Os sinais verticais da mesoderme e os sinais planares do lábio dorsal do blastoporo permitem a indução da ectoderme e a formação do tubo neural.

Fig.10 - Diferença entre os sinais verticais e os planares
Sinais verticais
Folistatina
Esta proteína é codificada por um RNAm maternal no ovo e é secretada e acumulada na zona marginal dorsal. Durante a gastrulação, esta acumula-se na mesoderme dorsal anterior.
Quando ela se associa à activina:
- activina + receptor - diferenciação ectodermica e inibição da diferenciação neural
- activina + folistatina - indução da diferenciação neural.
Noggin
A baixas concentrações ela induz a ovogénese e a fecundação, no entanto, a fortes concentrações induz a gastrulação na zona marginal dorsal e a migração.
Proteína Chordal
Quando associada às células do polo animal, dá as células do tipo neural.
Proteína Cerberus
Está presente na região dorsal durante a gastrulação.
Proteínas Frzb e Dickoff
São sintetizadas na mesoderme e numa expressão elevada vão provocar a ausência de estruturas ventrais, ou seja, não vão ter mais que as cabeças.

Fig.11 - Funcionamento da proteína Fzrb
Mesmo sem mesoderme, se metermos as proteínas Chordal e Noggin em contacto com a ectoderme, ela vai-se tornar neuroderme, bloqueando BMP4. No entanto, com a proteína BMP4 estas células vão dar epiderme, porque esta molécula inibe a diferenciação neural e então, a formação da neuroderme.

Fig.12 - Formação da neuroderme e epiderme
Sinais planares
Ácido retinóico
Participa na formação do eixo antero-posterior, promovendo a formação de estruturas neurais, troncais e posteriores.
Proteína Wint
Responsável por formar as partes posteriores; indução planar posterior.
FGFs
Participam no desenvolvimento de estruturas neurais posteriores.
Cerebrus + frizber + chordal + folistatina + noggin = activação neural da ectoderme dorsal
Ácido retinóico + FGFs + Wint = formação de estruturas posteriores ao longo do eixo antero-posterior

Fig.13 - Distribuição de todos os sinais verticais e planares
As cinco funções do centro organizador de Spemann:
- Controlo da inibição da gastrulação: iniciação da invaginação
- Produzir as células da mesoderme axial
- Dorsalisar a ectoderme: organização espaço-temporal da placa neural
- Induzir o tubo neural
- Dorsalisar a mesoderme da zona marginal
Formação e regionalização do sistema nervoso - NEURULAÇÃO
Na neurulação, dá-se a formação, a partir da ectoderme, do tubo neural, da epiderme e das células da crista neural, em que cada um dará órgãos ou tecidos diferentes.
Do tubo neural desenvolver-se-á o SNC (encéfalo e medula espinal), e da epiderme as unhas, pêlos, entre outros. As células da crista neural formam-se após a formação do tubo neural na região mais dorsal deste e darão estruturas como por exemplo os neurónios do SNP.

Fig.14 - Formação e fecho do tubo neural
A partir da ectoderme forma-se inicialmente a placa neural. Seguidamente as células da placa neural mudam de conformação e começam a encurvar para o interior, formando lateralmente as pregas neurais até que esta se juntam fechando o tubo neural e formando na sua parte mais dorsal as células da crista neural.
Antes do fecho completo do tubo neural, as pregas ao juntarem-se formam dois neuroporos: anterior e posterior.

Fig.15 - Localização dos neuroporos anterior e posterior
O neuroporo anterior fecha-se aos 24 dias após a fecundação e o neuroporo posterior apenas aos 26 dias.
A formação das pregas neurais e fecho do tubo neural dá-se devido a uma mudança de forma das células da ectoderme induzida pela notocorda.
Os sinais enviados pela notocorda induzem a um aumento da espessura e comprimento das células, passando estas a chamar-se células neuroepiteliais e formando a placa neural. Estas células apresentam agora uma forma cilíndrica com filamentos de actina no topo dos microtubulos, na parte mais dorsal das células.
Com a continuação da indução pela notocorda, estes filamentos de actina formam um anel apertando os microtubulos no topo fazendo com que estes se "abram" na parte inferior provocando um alargamento da célula em baixo e estreitamento em cima. As células ficam agora com forma trapezóide encurvando a placa.

Fig.16 - Indução das células da ectoderme pela notocorda
Na placa neural, as células que darão o tubo neural e as células que darão a epiderme possuem caderinas diferentes. Caderinas são moléculas de adesão celular pertencentes à família das CAM, que permitem a ligação com as células vizinhas. As células da placa neural possuem N-caderina e as células da futura epiderme E-caderina. Por isso as células neuroepiteliais ligar-se-ão entre elas após o encurvamento das mesmas, e as células que darão a futura epiderme também.
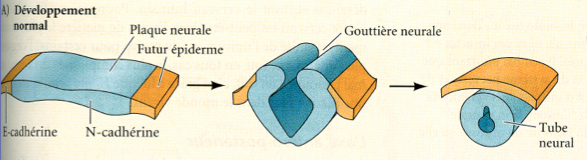
Fig.17 - Especificação das células
Após fecho do tubo neural, dá-se diferenciação do mesmo. O primeiro acontecimento que ocorre é a síntese das sub-populações de neurónios. Na parte superior existe uma parte designada por roof plate e na parte inferior uma designada por floor plate. A roof plate emite sinais BMPs que induzem a diferenciação das células neurais do neuroepitélio em neurónios, que são classificados em neurónios dorsais ou ventrais conforme a sua disposição ao longo do eixo dorso-ventral.

Fig.18 - Diferenciação do tubo neural
A ventralização do tubo neural é induzida pela floor plate (FP) e pela notocorda, que libertam um factor ventralizante que irá provocar a diferenciação das células ventrais do tubo em placa basal e neurónios motores.
O factor ventralizante libertado é a proteína Sonic Hedghog (SHH) e a diferenciação irá depender da concentração desta mesma proteína.

Fig.19 - Experiências que provam a expressão da proteína SHH
Por isso, colocando a placa neural em contacto com a FP ou directamente com a proteína SHH resulta igualmente na produção das estruturas ventrais. A baixa concentração de SHH provoca a ausência das estruturas ventrais.

Fig.20 - Diferenciação das células neurais do tubo neural consoante os sinais dorsalizantes ou ventralizantes
Formação das células da crista neural
As células da crista neural, localizadas na parte mais dorsal do tubo neural após o fecho deste, provêm das células situadas entre a ectoderme e a neuroectoderme (marcado a vermelho na Fig.21), situadas nas pregas neurais.

Fig.21 - Formação das células da crista neural
Estas células desagregam-se devido à paragem de expressão da N-caderina provocando uma perda de coesão celular e migração destas. Na migração destas, volta a haver expressão de N-caderina e estas agregam-se em gânglios.
As células da crista neural (CCN) são multipotentes, e por isso, durante e após a migração diferenciam-se formando três grandes tipos celulares:
- Células do SNP
- Células pigmentares
- Células de schwann dos nervos periféricos

Fig.22 - Diferenciação das células da crista neural
Existem vários tipos de células da crista neural:
- CCN da cabeça
- CCN du tronco
- CCN vagais e sacrais
- CCN cardíacas
Formação e regionalização dos somitos - SOMITOGÉNESE
Os somitos provêm da mesoderme, mais propriamente da mesoderme paraxial. Começam-se a formar imediatamente após a gastrulação, de maneira regular, da frente para trás.
No início existe a mesoderme paraxial sem qualquer organização, seguidamente dá-se epitelialização e as células começam a organizar-se.

Fig.23 - Evolução da mesoderme paraxial até aos somitos
- esclerotómos
- miotómos
- dermatomos

Fig.24 - Formação dos somitos
No fim da gastrulação dá-se:
1) Generação da mesoderme paraxial
2) Generação de um padrão de segmentação
3) Determinação de uma identidade axial
4) Compartimentação antero-posterior dos somitos
5) Formação de um somito epitélial
6) Polarização somítica
A organização segmentar dos somitos é a origem do padrão de segmentação do esqueleto axial e do sistema nervoso.
O esqueleto axial, a derme dorsal e a totalidade das fibras musculares (excepto da cabeça) são derivadas dos somitos.
Ao nível dos somitoméros, todas as células são totipotentes. Estas células sofrem um mecanismo de diferenciação que corresponde à origem do esclerotómo (cartilagem das vértebras e costelas), do miotomo (fibras musculares dos membros) e do dermatomo (derme dorsal).
Nos somitoméros, a parte mais próxima do tubo neural vai dar os músculos epaxiais, e na parte oposta vão se formar os músculos hipoaxiais.
A ordem de aparição dos somitos ao longo do eixo antero-posterior (da cabeça até à cauda) é determinada durante a gastrulação, pelos genes homeóticos.
Existem sinais de ventralização (SHH) resultantes da corda que vão induzir as células ventrais - esclerotomo (cartilagem). A epiderme vai induzir as células que vão formar o dermatomo (Wint) - sinais dorsalizantes.
Existem também sinais provenientes das lâminas laterais (BMP4) que vão induzir a formação do miotomo (músculos hipoaxiais).
Sem comentários:
Enviar um comentário